
“There is nothing more deceptive than an obvious fact.”
Arthur Conan Doyle, The Boscombe Valley Mystery
I have watched or been a student in a lot of biology classes over the years. I sometimes think we take a lot for granted when we teach students. Not only in biology but in many of the STEM fields. We have the advantage of teaching science on the shoulders of all the greats that came before us. Sometimes I think we forget how long it took to answer questions and just how smart the people that figure them out were. Also, we forget how fast things change, in biology we have something called The Central Dogma. Simply it states that DNA goes to RNA goes to protein. It’s as simple as that; we know proteins are not made directly from DNA and RNA is not made from proteins.
The funny thing about The Central Dogma’s place in modern biology is that it’s relatively new. We’ve been studying biology for a long time; Van Leeuwenhoek discovered single-cell organisms in 1670, Hooke coined the term cell in 1665. Macromolecules came later; Proteins in 1838, DNA in 1869, and RNA between 1890 and 1950, RNA was initially thought to be the same as DNA. However, we didn’t know whether DNA or proteins were the sources of genetic inheritance until 1952. We didn’t know the structure of DNA until 1953. Meselson and Stahl published the proof of semi-conservative replication of DNA in 1958.
In 2018 most of The Central Dogma is less than 70 years old. There are a substantial number of people alive that are older than The Central Dogma. This information is only old because of the speed at which biology has been progressing over the last century.
When teaching facts In STEM education we often run into a severe problem, students can often give us the “correct” answer on a test. However, if you dig a little deeper, they don’t understand what that answer means.
I have often thought that teaching biology (or any STEM field) through an understanding of the foundational experiments would help students understand the facts. Imagine going through these experiments; What was the question?, Why did they do this?, Why didn’t they do that?, What do the results show?, and What do they do next?. Teaching these experiments to students would explain not only what we know but why we know it.
Let’s look at a couple of examples. We know that DNA is the molecule responsible for genetic inheritance. How do we know? For many years scientists thought proteins had to be the source of genetic inheritance because DNA was just too simple. In 1952 Alfred Hershey and Martha Chase conducted an experiment that provided some of the most persuasive evidence that DNA was the source of genetic inheritance.
Hershey and Chase use T2 bacteriophage for their experiment, T2 phage reproduced by infecting a bacterial cell. The bacterial cell produces new phage that would be released when the cell lysed. While the mechanism of T2 phage reproductions was not known, the process required the transfer of “genetic material” from the phage to the bacteria. The T2 phage is composed of two components a protein shell and DNA core. The researchers needed to determine what part of the T2 phage entered the bacterial cell.
The researchers needed a way to label proteins and DNA independently of each other. Two atoms helped sulfur and phosphorus. Proteins use sulfur while DNA does not. DNA uses phosphorus while proteins do not. They grew phage with radioactive sulfur or radioactive phosphorus. These radioactive phages infected cells, after infection, the phage and cells were separated, and the location of the radioactivity was determined.
They found the radioactive DNA was always with the bacteria (Figure 1B) while none of the radioactive protein was with the bacteria (Figure 1A). They also showed that radioactive DNA could get incorporated into the bacterial DNA. While other scientists conducted additional experiments, this experiment showed it was the DNA, which carried the genetic information.

Your students can probably (we hope) tell you that DNA replicates semiconservatively. However, if you asked them to prove semiconservative replication of DNA, could they do it? Without looking up the Meselson and Stahl experiment. In the late 1950s when Matthew Meselson and Franklin Stahl conducted their research, we already knew the structure of DNA. It was immediately clear from the structure that DNA could serve as a template for its replication.
Early on there were three competing models for DNA replication; conservative replication (Figure 2A), semiconservative replication (Figure 2B), and dispersive replication (Figure 2C). The differences in these models can be described based on where the new and old DNA strands are after replication. In conservative replication after one round, you end up with one DNA molecule composed entirely of new DNA and one molecule composed entirely of old DNA. After two rounds of replication, you now have three new DNA molecules and one old DNA molecule. In semiconservative replication after the first round, you get two molecules that both contain one new and one old strand of DNA. After two rounds you get two molecules composed entirely of new DNA and two molecules composed of one new and one old strand. In disrupted replication, the DNA molecule was cut every ten base pairs on alternating strands, and then new DNA would fill in the gaps. After one round you get molecules that are 50-50 old versus new DNA. After two rounds of replication, you get four strands that would have somewhere between 50-50 and 75-25 new versus old DNA.
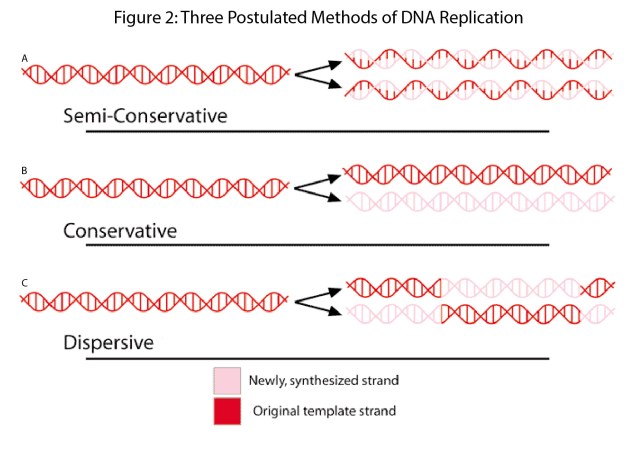
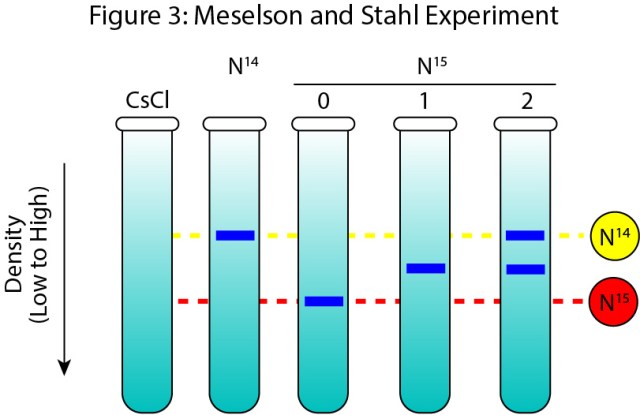
The beauty of these models is that if you can follow the new and the old DNA you can distinguish between all models. Meselson and Stahl marked new and old DNA with nitrogen isotopes specifically N14 and N15 these isotopes differ by one neutron. Which turns out is enough to separate DNA by density in a cesium chloride gradient.
They grew bacteria on media which contained N15 then allowed the cells to grow on media containing N14 for 0, 1, or 2 cycles of replication. The DNA was then isolated from the cells and density was used to separate the DNA molecules. After zero rounds of replication, there was a single band lower than cells grown only on N14 (Figure 3 N15 0). After one round of DNA replication, there was a single band between the N15 and N14 bands (Figure 3 N15 1). This result ruled out conservative replication since conservative replication should have produced one heavy (N15) and one light (N14) band. However, both semiconservative and disrupted replication should produce 50-50 molecules at round one. After two rounds of replication, we get two band’s one at the 50-50 spot the second at the light (N14) position (Figure 3 N15 3). This position of bands is what you’d expect from semiconservative replication but not dispersive replication. Dispersive replication would have produced a band between the 50-50 and the N14 band. Therefore, DNA replicated semiconservative.
Even if you don’t need this experiment to teach your students how semiconservative replication works, the Meselson and Stahl experiment is often referred to as one of the most elegant experiments ever conducted in biology and is worth studying to learning experimental design.
Up until these experiments were conducted the information that we teach as clear and obvious facts was up for debate. While we probably can’t go over every single fundamental experiment in enough details, so our students understand them, because of the total amount of material we need to cover, foundational experiments can be useful. If there’s a topic that your students are having trouble grasping maybe take the students through the experiments that demonstrated the facts. Perhaps the solution is a one-credit recitation that covers the experiments in conjunction with the lector. That might solve all our problems (shakes head ruefully). One last thought, if students are having trouble grasping that clear and obvious fact maybe stop and ask if it is clear and obvious?
Thanks for listening to my musings
The teaching cyborg

{kind=link}
{kind=link}
{kind=link}